Special
Mechanism of Flapping Flight in Hummingbirds
Author: Hiroto Tanaka
*This article is an English version of a review contributed to the book “The Most Beautiful Hummingbird Encyclopedia in the World” (written and edited by Hiroya Mizuguchi, Seibundo Shinkosha, 2025, ISBN 9784416723333). AI tool was used for translation.

Photo: Hiroto Tanaka
Ecology of Flight
Hummingbirds, as their name suggests, are small birds that hover by flapping their wings rapidly, much like bees, while feeding on floral nectar. Their English name, “hummingbird,” is derived from the humming sound generated by their wingbeats. They belong to the family Trochilidae within the order Apodiformes (sometimes classified under Caprimulgiformes), comprising 366 known species [1]. Their distribution ranges from South America to North America and Canada, with some species undertaking long-distance migrations within the Americas.
A defining feature of hummingbird flight is hovering, in which the body is maintained stationary in midair through rapid wing flapping. This enables them to feed on nectar while hovering. This mode of flight contrasts with that of swifts (family Apodidae), which are adapted for gliding flight with minimal flapping and primarily capture airborne insects.
In addition to hovering, hummingbirds can rapidly alter their position and body orientation, or execute abrupt reversals in flight. Such exceptional flight capabilities are of interest not only to biologists but also to engineers. Although nectar constitutes their primary food source, hummingbirds frequently consume insects and spiders. They capture prey both from surfaces such as leaves and nests, and directly in flight [2] (Stiles, 1995). Their high maneuverability plays a crucial role in such aerial predation.
Hummingbirds are also capable of long-distance migration. Species that migrate seasonally overwinter in warm regions such as northern South America, Central America, and the southern United States, and spend the breeding season in North and Central America. Their migration patterns can be visualized using online platforms that compile and map observational data [3] (Hummingbird Central).
Species known to migrate distances exceeding 1000 km include the Ruby-throated Hummingbird, Green-backed Firecrown, Ruby-topaz Hummingbird, Black Jacobin, Planalto Hermit, and Black-throated Mango [4] (Rappole and Schuchmann, 2003).
Among these, the Ruby-throated Hummingbird performs a remarkable non-stop flight across the Gulf of Mexico, traveling from the Yucatán Peninsula to the southern United States. This journey spans approximately 800–1000 km and may last up to 20 hours. By utilizing seasonal tailwinds, flight speeds are estimated to reach 40–80 km/h. Prior to migration, individuals accumulate energy reserves, increasing their body mass from approximately 3 g to 5 g through fat deposition of about 2 g [5] (Hargrove, 2005).
Morphology as an Aircraft
The mechanisms and characteristics of flight are influenced by body mass, wing size, and wing planform.
With respect to body mass, hummingbirds constitute the smallest and lightest group among birds. The lightest species is the male Bee Hummingbird, with an average mass of 1.74 g [6] (Lawrie et al., 2021). As its name implies, it is comparable in size to a bee. Most species weigh less than 6 g, while even larger species typically remain below 12 g. An exception is the Giant Hummingbird, which reaches approximately 20 g [7] (Fernandez et al., 2011).
Wing size increases with body mass, whereas wingbeat frequency decreases. For example, the Rufous Hummingbird and Black-chinned Hummingbird, each weighing approximately 3 g, exhibit wing lengths of about 42 mm and 47 mm, with wingbeat frequencies of approximately 52 Hz and 51 Hz, respectively (Footnote 1). In contrast, larger species such as Rivoli’s Hummingbird (~7 g) and Blue-throated Hummingbird (~8 g) have wing lengths of approximately 79 mm and 85 mm, and lower wingbeat frequencies of about 24 Hz and 23 Hz, respectively [8] (Chai and Millard, 1997). The Giant Hummingbird (~20 g) exhibits a wing length of approximately 140 mm and a wingbeat frequency of about 13 Hz [9] (Fernandez, 2010). Overall, hummingbirds are characterized by high-frequency flapping of relatively small wings compared with other birds.
This high-frequency flapping is enabled by the specialized musculoskeletal structure of the wing. Because both the humerus and forearm are extremely short, extension and flexion of the wing are limited, allowing the wings to remain extended and flap at high speed, similar to insect wings [10, 11] (Tobalske et al., 2007; Warrick et al., 2012).
The wing mass accounts for approximately 5–7% of total body mass, while the pectoral muscles responsible for flapping constitute about 30% [7] (Chai and Millard, 1997). The mass ratio of downstroke to upstroke muscles is approximately 1.7:1 in hummingbirds, whereas in other birds it is typically around 10:1, indicating a relatively greater development of upstroke musculature in hummingbirds [12] (Deeming, 2023). This difference is closely related to their distinct flight mechanics.
The wing planform is characterized by the aspect ratio, defined as the ratio of wing length to mean chord length (wing area divided by wing length). Larger aspect ratios indicate long, slender wings. Hummingbird wings exhibit aspect ratios between 3 and 4.5, indicating relatively slender shapes [13] (Kruyt et al., 2014). High aspect ratio wings generally achieve higher lift-to-drag ratios, enhancing aerodynamic efficiency. The concepts of lift and drag are discussed in the following section (Footnote 2).
Footnotes
(1) In this article, wing length is defined as the distance from the wing root to the wing tip. In contrast, in avian morphological terminology, “wing length” refers to the length of the folded wing, that is, the distance from the wrist joint to the tip of the primary feathers.
(2) In aerodynamics of aircraft, the aspect ratio is defined using the total wing area of both wings and the distance between the wingtips (i.e., the wingspan). Therefore, for hummingbirds, the aerodynamic aspect ratio corresponds to approximately twice the value defined above, yielding a range of about 6 to 9.
Aerodynamics of Hovering
During hovering, hummingbirds orient their body axis in a slightly inclined posture and flap their wings obliquely downward from a rear-upward to a front-downward direction (downstroke from dorsal to ventral side), and then upward from front-downward to rear-upward (upstroke from ventral to dorsal side) [10] (Tobalske et al., 2007).
The aerodynamic forces generated by the wings can be decomposed into lift and drag (Fig. 1). Lift is the component perpendicular to the relative airflow, while drag is the component parallel to it. When hummingbirds flap their wings approximately horizontally, the relative airflow is also nearly horizontal, and thus the lift component is directed almost vertically upward during both downstroke and upstroke.
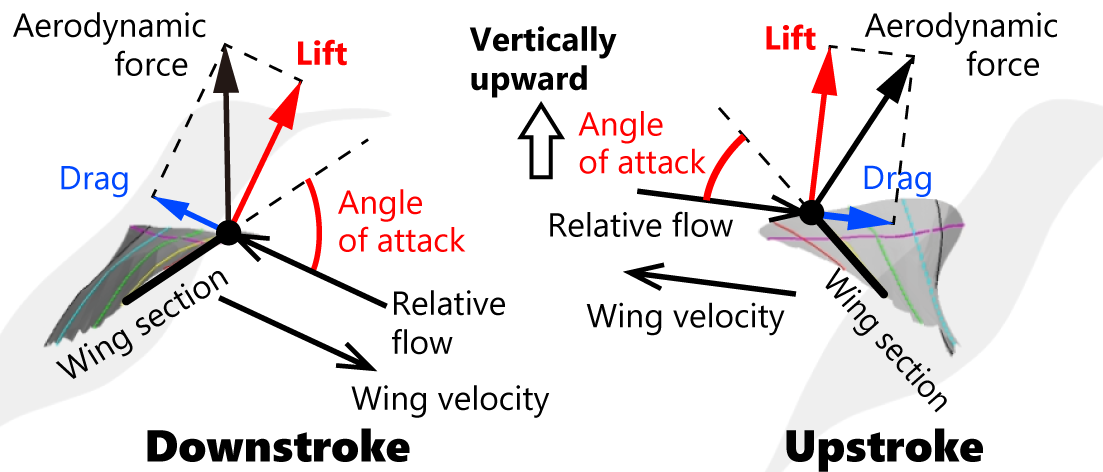
Figure 1. Aerodynamic forces acting on a flapping wing of a hummingbird during downstroke and upstroke.
Thus, during hovering, both the downstroke and upstroke contribute to supporting body weight primarily through lift generation. In contrast, other small birds performing brief hovering expand their wings during the downstroke but fold them during the upstroke by bending the elbow and wrist joints. Consequently, these birds rely mainly on the downstroke, whereas the upstroke contributes little. This difference in flight mode is reflected in the previously described distribution of pectoral muscle mass.
Lift generally increases with the angle of attack, defined as the angle between the wing and the relative airflow. However, if the angle of attack becomes excessively large, flow separation occurs, resulting in a reduction of lift and an increase in drag. This phenomenon is known as stall (Fig. 2, upper left). To maintain an appropriate angle of attack and avoid stall, hummingbird wings undergo torsional deformation in opposite directions during downstroke and upstroke [14, 15] (Maeda et al., 2017; Song et al., 2014). This twisting is likely facilitated by passive deformation of the flight feathers.
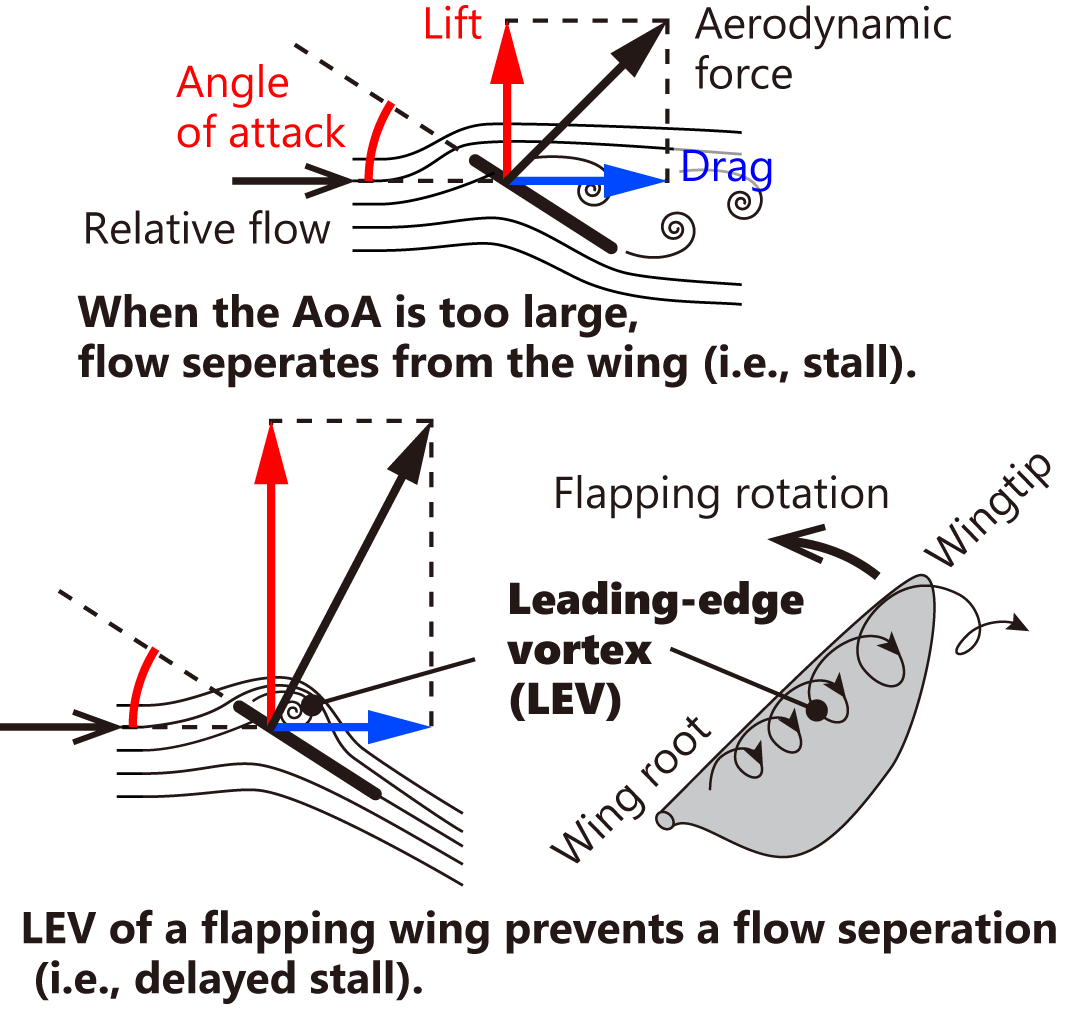
Figure 2. Flow separation from the wing and its suppression by leading-edge vortices in flapping wings.
The angle of attack at the mid-span of the wing is approximately 40° during mid-downstroke and 30° during mid-upstroke [14, 15] (Maeda et al., 2017; Song et al., 2014). These values are sufficiently large to induce flow separation in conventional aircraft wings. However, hummingbirds utilize the leading-edge vortex (LEV), a characteristic feature of flapping wings with small and thin profiles, to suppress flow separation and sustain high lift.
In fact, as wing size decreases, the relative importance of viscous forces increases compared with inertial forces, resulting in more prominent vortex structures. Furthermore, thin wings with sharp leading edges promote the formation of strong vortices at high angles of attack. While a stationary wing experiences only a short duration of vortex attachment before separation occurs, a rotating (flapping) wing maintains a spiral leading-edge vortex attached over a longer duration, preventing flow separation and sustaining high lift (Fig. 2, lower right) [16] (Sane, 2003).
Numerical simulations of a hovering Ruby-throated Hummingbird (body mass 3.4 g, wing length 45 mm, wingbeat frequency 42 Hz) revealed that a leading-edge vortex remains attached to the wing surface during downstroke, generating substantial lift. During upstroke, although less pronounced, a leading-edge vortex is also formed and contributes to lift generation. As a result, the vertical component of aerodynamic force during downstroke is approximately 2.5 times that during upstroke. This difference arises from factors such as higher flapping velocity and angle of attack during downstroke, slightly larger effective wing area, and the upward component of drag during downstroke [15] (Song et al., 2014).
Diverse and Agile Flight
Hummingbirds are not only proficient at hovering but also capable of highly diverse and agile flight by modulating wing kinematics. Several representative studies illustrating these capabilities are introduced below.
Experiments have examined how much load hummingbirds can lift while hovering by attaching weights to their bodies [8] (Chai and Millard, 1997). Larger species such as Rivoli’s Hummingbird (~7 g) and Blue-throated Hummingbird (~8 g) were able to lift loads up to approximately twice their body weight for short durations (~0.4 s).
In smaller species such as the Rufous Hummingbird and Black-chinned Hummingbird (~3 g), individuals were able to lift loads approximately equal to their body weight for about 0.6 s. During maximum load conditions, wingbeat frequency increased by approximately 17–33%, and stroke amplitude increased by approximately 13–29% compared with unloaded conditions. These results indicate that hummingbirds can transiently generate aerodynamic forces equivalent to three to four times their body weight by increasing flapping frequency and amplitude, enabling rapid upward acceleration.
The maximum forward flight speed has been measured in wind tunnel experiments using Ruby-throated Hummingbirds (~4 g) [17] (Chai and Dudley, 1999). The average maximum speed was 14.4 m/s (52 km/h). During molting, when flight feathers are replaced, the maximum speed decreased to 12.4 m/s (45 km/h).
Similar experiments on the Rufous Hummingbird [10] (Tobalske et al., 2007) showed maximum speeds of approximately 12 m/s (43 km/h). In hovering, the body axis is inclined forward by about 50° from the horizontal, whereas at higher speeds the body becomes more horizontal, reaching approximately 13° at maximum speed. Conversely, the wingtip trajectory, viewed laterally, shifts from about 15° above horizontal during hovering to approximately 68° at maximum speed (Fig. 3). Although hummingbird wing motion is often described as a figure-eight pattern, it does not always form a clear figure-eight trajectory.
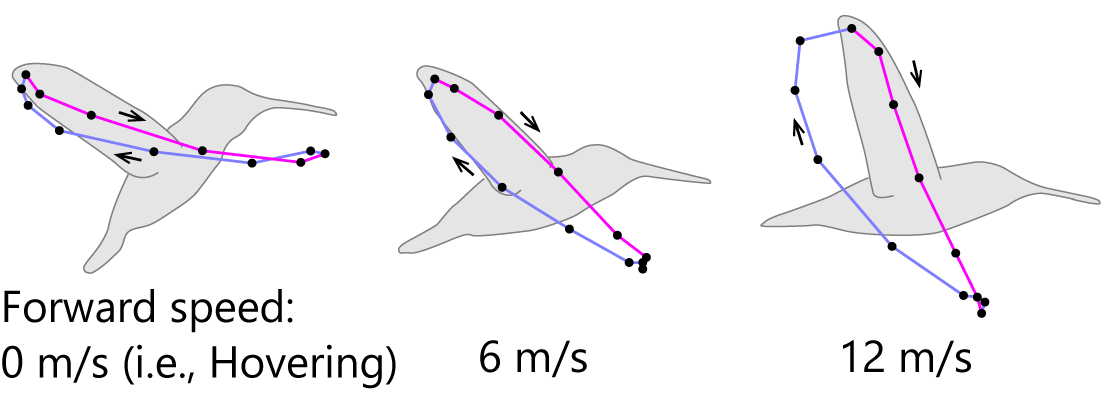
Figure 3. Wingtip trajectory of the Rufous Hummingbird viewed laterally (modified from [10]).
Hummingbirds are also capable of backward flight. In wind tunnel experiments, Anna’s Hummingbirds (~4.6 g) were induced to fly backward by applying airflow from behind while hovering and feeding [18] (Sapir and Dudley, 2012). The maximum backward flight speed was 4.5 m/s (16 km/h). In this condition, the body axis becomes nearly upright (71° from horizontal), and the flapping plane shifts accordingly. Thus, forward and backward flight are achieved through adjustments of body orientation and wing kinematics.
Hummingbirds can also cope with turbulent airflow. When airflow disturbances were generated by placing a cylinder upstream in a wind tunnel, Anna’s Hummingbirds maintained their position by asymmetrically adjusting wing motions and utilizing tail feathers for stabilization [19] (Ortega-Jimenez et al., 2014).
Furthermore, hovering Anna’s Hummingbirds can pass through narrow apertures by laterally translating their body while reducing wingbeat amplitude. When the aperture becomes even smaller, they transition to forward flight, tilt their body forward, retract their wings, and pass through the opening without flapping, similar to a projectile [20] (Badger et al., 2023).
When a hovering hummingbird feeding on nectar is startled by an approaching object from the front, it performs a rapid escape maneuver by reversing direction and flying backward. This maneuver is achieved within approximately five wingbeats through a pitch-up followed by a roll motion [21] (Cheng et al., 2016). The pitch-up rotates the body backward within approximately two wingbeats (~0.06 s), followed by a roll over the next three wingbeats (~0.09 s), restoring the ventral side downward and completing reorientation. The entire maneuver occurs within approximately 0.15 s.
The reaction time to visual stimuli is also remarkably short. In experiments involving four species (Rivoli’s Hummingbird, Blue-throated Hummingbird, Black-chinned Hummingbird, and Broad-billed Hummingbird), three species exhibited response times shorter than 0.03 s, which is faster than the typical reaction time of insects (~0.05 s). The remaining species showed a response time of approximately 0.05 s, comparable to that of insects. Such rapid response times contribute significantly to the agility of hummingbird flight.
In summary, hummingbirds achieve hovering and highly agile flight by flapping their small wings at high frequency while dynamically modulating wing motion. The limits of their flight performance remain incompletely understood, and further studies are ongoing. This article has emphasized explanations based on reliable data and cited numerous research papers. Some of these references are freely accessible online, and some include video materials. Such references are indicated as (free/video) in the reference list. Readers interested in this topic are encouraged to consult these papers and explore the field of hummingbird research in greater depth. (Hiroto Tanaka, PhD in Information Science and Engineering)
References
[1] Gill, F., Donsker, D. and Rasmussen, P., eds. (2024). “Hummingbirds” IOC World Bird List. V14.2.
https://www.worldbirdnames.org/ (Website)
[2] Stiles, F. G. (1995). Behavioral, Ecological and Morphological Correlates of Foraging for Arthropods by the Hummingbirds of a Tropical Wet Forest. The Condor 97, 853–878.
https://doi.org/10.2307/1369527
[3] Hummingbird Central.
https://www.hummingbirdcentral.com/hummingbird-migration.htm (Website)
[4] Rappole, J. H. and Schuchmann, K.-L. (2003). Ecology and Evolution of Hummingbird Population Movements and Migration.
https://doi.org/10.1007/978-3-662-05957-9_3
[5] Hargrove, J. L. (2005). Adipose energy stores, physical work, and the metabolic syndrome: lessons from hummingbirds. Nutrition Journal 4.
https://doi.org/10.1186/1475-2891-4-36 (Free)
[6] Lawrie, Y., Shannon, T. J., Kirkconnell, A., Clark, C. J., Collinson, J. M. and Kirwan, G. M. (2021). Systematic placement of the bee hummingbird (Mellisuga helenae) (Aves: Trochilidae) and potential consequences for nomenclature of the Mellisugini. Ornitologia Neotropical 32, 45–50.
https://doi.org/10.58843/ornneo.v32i1.947
[7] Fernandez, M. J., Dudley, R. and Bozinovic, F. (2011). Comparative Energetics of the Giant Hummingbird (Patagona gigas). Physiological and Biochemical Zoology 84, 333–340.
https://doi.org/10.1086/660084
[8] Chai, P. and Millard, D. (1997). Flight and size constraints: Hovering performance of large hummingbirds under maximal loading. Journal of Experimental Biology 200, 2757–2763.
https://doi.org/10.1242/jeb.200.21.2757 (Free)
[9] Fernandez, M. J. (2010). Flight Performance and Comparative Energetics of the Giant Andean Hummingbird, PhD thesis, UC Berkeley.
https://escholarship.org/ (Free)
[10] Tobalske, B. W., Warrick, D. R., Clark, C. J., Powers, D. R., Hedrick, T. L., Hyder, G. A. and Biewener, A. A. (2007). Three-dimensional kinematics of hummingbird flight. Journal of Experimental Biology 210, 2368–2382.
https://doi.org/10.1242/jeb.005686 (Free)
[11] Warrick, D., Hedrick, T., Fernández, M. J., Tobalske, B. and Biewener, A. (2012). Hummingbird flight. Current Biology 22, R472–R477.
https://doi.org/10.1016/j.cub.2012.04.057 (Free)
[12] Deeming, D. C. (2023). Allometry of the pectoral flight muscles in birds. Journal of Zoology 319, 264–273.
https://doi.org/10.1111/jzo.13043 (Free)
[13] Kruyt, J. W., Quicazán-Rubio, E. M., van Heijst, G. F., Altshuler, D. L. and Lentink, D. (2014). Hummingbird wing efficacy depends on aspect ratio. Journal of the Royal Society Interface 11.
https://doi.org/10.1098/rsif.2014.0585 (Free)
[14] Maeda, M., Nakata, T., Kitamura, I., Tanaka, H. and Liu, H. (2017). Quantifying the dynamic wing morphing of hovering hummingbird. Royal Society Open Science 4.
https://doi.org/10.1098/rsos.170307 (Free & Movies)
[15] Song, J., Luo, H. and Hedrick, T. L. (2014). Three-dimensional flow and lift characteristics of a hovering hummingbird. Journal of the Royal Society Interface 11.
https://doi.org/10.1098/rsif.2014.0541 (Free & Movies)
[16] Sane, S. P. (2003). The aerodynamics of insect flight. Journal of Experimental Biology 206, 4191–4208.
https://doi.org/10.1242/jeb.00663 (Free)
[17] Chai, P. and Dudley, R. (1999). Maximum flight performance of hummingbirds. American Naturalist 153, 398–411.
https://doi.org/10.1086/303179
[18] Sapir, N. and Dudley, R. (2012). Backward flight in hummingbirds. Journal of Experimental Biology 215, 3603–3611.
https://doi.org/10.1242/jeb.073114 (Free & Movies)
[19] Ortega-Jimenez, V. M., Sapir, N., Wolf, M., Variano, E. A. and Dudley, R. (2014). Into turbulent air. Proc. Royal Society B 281.
https://doi.org/10.1098/rspb.2014.0180 (Free & Movies)
[20] Badger, M. A., McClain, K., Smiley, A., Ye, J. and Dudley, R. (2023). Sideways maneuvers in hummingbirds. Journal of Experimental Biology 226.
https://doi.org/10.1242/jeb.245643 (Free & Movies)
[21] Cheng, B., Tobalske, B. W., Powers, D. R., Hedrick, T. L., Wethington, S. M., Chiu, G. T. C. and Deng, X. Y. (2016). Flight mechanics of escape manoeuvres. Journal of Experimental Biology 219, 3518–3531.
https://doi.org/10.1242/jeb.137539 (Free)