Special
ハチドリの羽ばたき飛行のメカニズム
著:田中 博人
*本稿は、書籍「世界で一番美しいハチドリ図鑑」(編著:水口博也, 誠文堂新光社, 2025)に寄稿した記事を、編著者の了承のうえ転載したものです。

Photo: Hiroto Tanaka
飛行の生態
ハチドリは、その名の通りハチのように小さく羽ばたいて飛び、花の蜜を吸う。英語名はハミングバードhummingbirdで、羽ばたき音がブーンと鳴る(humming)ことに由来する。鳥綱アマツバメ目(ヨタカ目とする説もあり)ハチドリ科に分類され、既知の種数は366種を数える[1]。南米から北米およびカナダにかけて生息し、南北アメリカ内で長距離の渡りをする種もある。
ハチドリを特徴づける飛び方は、高速の羽ばたきによって空中で体を静止させるホバリングである。ホバリングしながら花の蜜を吸うこともできる。これは、同じアマツバメ目のアマツバメ科の鳥が、羽ばたかない滑空飛行を得意として主に空中の昆虫を捕食するのとは対照的である。
ハチドリはホバリングできるだけでなく、その位置や姿勢を瞬時に変えたり、急反転して飛び去ることもできる。このような優れた飛行能力は、生物学者だけでなく、私のような工学者も魅了する。ハチドリは花蜜を主食とするが、 (ハチドリはそもそも昆虫やクモを一定の割合で食べるようです。必要があれば帰国して論文探します。)昆虫やクモも頻繁に食べる。葉や巣についている虫だけでなく、飛行中の虫も、空中で捕食する[2](Stiles, 1995)。その際には優れた俊敏性が役立つ。
ハチドリは長距離の移動も可能である。季節毎に渡りをする種は、冬期は温暖な南米北部、中米、北米南部ですごし、春から秋は北中米内の各繁殖地ですごす。渡りの様子は、さまざまなハチドリの目撃情報を地図上で可視化しているウェブサイトで確認できる[3](Hummingbird Central)。
1000 km以上の長距離を移動する種として、ノドアカハチドリRuby-throated Hummingbird、ベニイタダキハチドリGreen-backed Firecrown、ルビートパーズハチドリRuby-topaz Hummingbird、クロハチドリ Black Jacobin、バライロユミハチドリPlanalto Hermit、ムナグロマンゴーハチドリBlack-throated Mango が知られている[4](Rappole and Schuchmann, 2003)。
なかでもノドアカハチドリは、メキシコのユカタン半島からアメリカ南部に向かって、メキシコ湾の海上を無着陸で一気に飛び越える。その距離は約800 kmから1000 kmにおよび、飛行時間は20時間にもなり、南から北への季節風も利用して飛行速度は時速約40から80kmになると推測される。渡り前にはエネルギーを蓄えるために、脂肪が約2gも増量して体重が5gになる[5](Hargrove, 2005)。
飛行体としての形態
飛行のメカニズムや特性は、飛行体の重量、翼の大きさ、翼の平面形状によって変わる。
まず体重については、ハチドリは鳥のなかでもっとも軽くて小さいグループである。もっとも軽いのはマメハチドリBee Hummingbirdの雄で、平均1.74gという記録がある[6](Lawrie et al., 2021)。その英名どおり、まさに「ハチ」ドリである。ほとんどの種は体重が6g以下であり、それ以外も12g以下である。例外がオオハチドリGiant Hummingbird で、体重は約20gもある[7](Fernandez et al., 2011)。
翼は、体重が軽いほど小さく、体重が重いほど大きい。逆に、一秒間の羽ばたき回数は、体重が軽いほど多く、体重が重いと少ない。たとえば、体重約3 gと軽いアカフトオハチドリRufous Hummingbird とノドグロハチドリBlack-chinned Hummingbird は、翼長と羽ばたき回数が、それぞれ約42mmと毎秒約52回および約47mmと毎秒約51回である(脚注1)。一方、ハチドリとしては大型で体重約7 gと重いアオノドハチドリRivoli’s Hummingbird と約8 gのルリノドシロメジリハチドリBlue-throated Hummingbirdは、翼長と羽ばたき回数はそれぞれ約79 mmと毎秒約24回および約85 mmと毎秒約23回である[8](Chai and Millard, 1997)。さらに体重約20 gのオオハチドリだと、翼長は約140 mm、羽ばたき回数は毎秒約13回である[9](Fernandez, 2010)。いずれにせよ、ハチドリは他の鳥にくらべて、小さな翼を高速で羽ばたかせる。
この速い羽ばたきを可能にするのが、ハチドリの翼の筋骨格である。上腕と下腕が非常に短いため腕の曲げ伸ばしがほとんどできず、まるで昆虫のように翼を広げたまま高速で羽ばたく[10, 11](Tobalske et al., 2007; Warrick et al., 2012)。
また、ハチドリの翼の重さは体重の約5%から7%で、翼を羽ばたかせる胸部の筋肉は体重の約30%を占める[7](Chai and Millard, 1997)。また胸部の筋肉のうち、打ち下ろし筋と打ち上げ筋の重量比は約1.7対1だが、他の鳥では10対1(打ち下ろし筋の方が極端に多い)にもなり、ハチドリでは打ち上げ筋の割合が他の鳥より多いことがわかる[12](Deeming, 2023)。ハチドリと他の鳥のこの筋肉配分の違いは、後述する飛び方の違いに関係する。
さて、翼の平面形状はアスペクト比で特徴づけられる。アスペクト比とは、平均翼弦長(1つの翼の面積を翼長で割った値)に対する翼長の比で表すことができ、値が大きいほど横に細長いことを意味する。ハチドリの翼のアスペクト比は3から4.5と大きく、細長い翼と言える[13](Kruyt et al., 2014)。アスペクト比が大きい翼は、基本的に抗力に対する揚力の比(揚抗比)が大きく、飛行に有利である。この揚力と抗力については次節で説明する(脚注2)。
脚注
(1)本稿では翼の根元から翼端まで長さとした。一方、鳥の形態用語における「翼長」とは、本稿の翼長とは異なり、翼をたたんだときの長さ、つまり手首から翼端=初列風切羽の端までの長さを指す。
(2)航空力学用語の「アスペクト比」は、左右2つの翼を合わせた翼面積と、左右の翼端間の距離、すなわち(翼幅(鳥の形態用語では翼開長)を用いてアスペクト比を算出する。ハチドリの場合、航空力学用語におけるアスペクト比は、先述の値を2倍にした6から9となる
ホバリングの空気力学
ハチドリがホバリングするとき、体軸を前傾気味に立て、翼を後上方から前下方に斜め下に打ち下ろし(背側から腹側への羽ばたき)、前下方から後上方に打ち上げる(腹側から背側への羽ばたき)[10] (Tobalske et al., 2007)。
翼が発生する空気力は、揚力と抗力に成分分解できる(図1)。揚力は、相対流速に垂直な方向の成分で、抗力は相対流速方向の成分である。ハチドリがほぼ水平に羽ばたくと、相対流速の方向もほぼ水平になるので、揚力は打ち下ろしと打ち上げのどちらでもほぼ鉛直上向きになる。
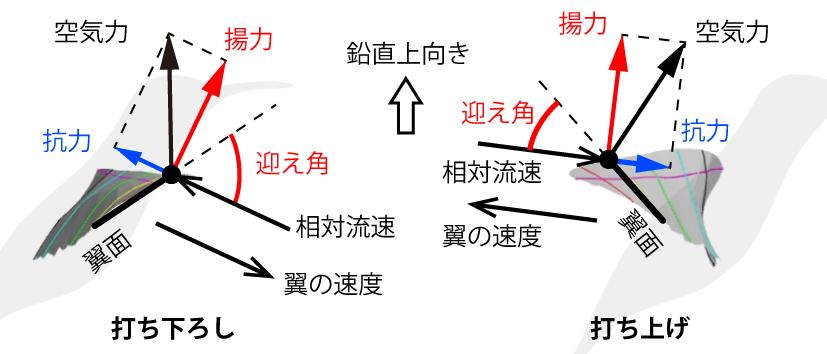
図1 ハチドリの羽ばたき翼の、打ち下ろし中と打ち上げ中と空気力
つまり、ハチドリのホバリングでは、打ち下ろしと打ち上げの両方が、主に揚力で体重を支える。一方、他の小鳥が短時間のホバリングをするときは、打ち下ろしでは翼を大きく広げるが、打ち上げでは翼を肘と手首で曲げて小さくたたむ。つまり、他の小鳥は打ち下ろしを利用するが、打ち上げはあまり利用しない。この飛び方の違いが、先述の胸部筋肉の質量比に表れている。
なお揚力は、翼と相対流速のなす角、すなわち迎え角が大きいほど比例して大きくなる。しかし、迎え角が大きすぎると、翼から流れが剥がれてしまい、揚力が減少して抗力が大きくなってしまう。これを航空力学では「失速」と呼ぶ(図2左上)。ハチドリでは失速しない適度な迎え角を保つために、翼は打ち下ろしと打ち上げでそれぞれ逆方向にねじれる[14, 15](Maeda et al., 2017; Song et al., 2014)。この翼のねじれには、翼を構成する風切羽の受動的な変形が寄与していると考えられる。
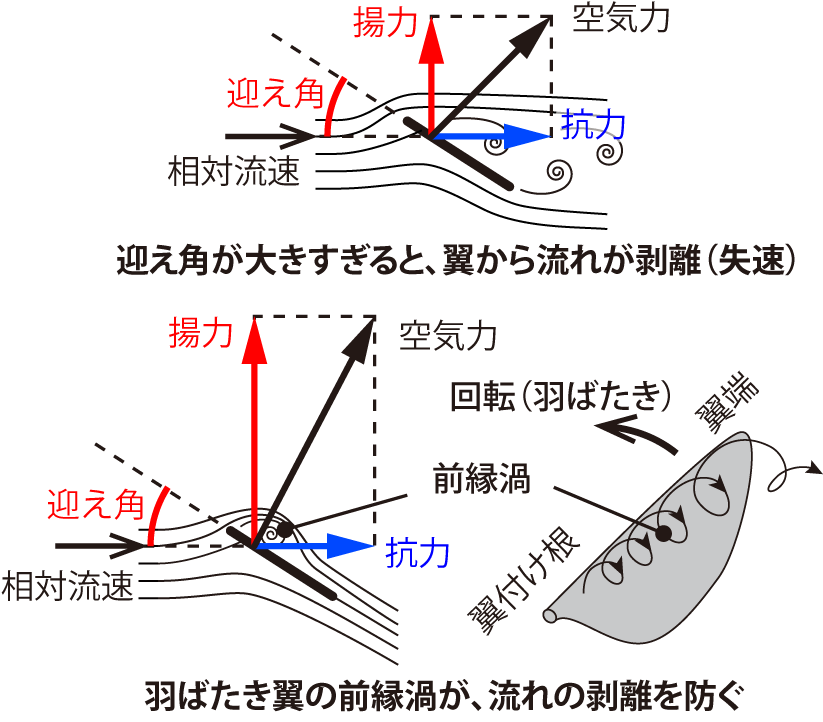
図2 翼から流れの剥離と、羽ばたき翼の前縁渦による流れの剥離防止
翼中央での迎え角の大きさは、打ち下ろし中期で約40°、打ち上げ中期で約30°である[14, 15](Maeda et al., 2017; Song et al., 2014)。これは、旅客機の翼ならば流れが剥離してしまうような大きな迎え角である。しかしハチドリは小さく薄い翼の羽ばたきに特有な「前縁渦」の効果を利用して、流れの剥離を抑えて大きな揚力を維持する。
じつは翼が小さいほど、空気の粘性が空気の慣性に対して相対的に強くなり、翼が形成する渦は相対的に大きく顕著になる。さらに、翼が薄くて前縁が鋭いと、大きな迎え角の時に前縁が強い渦を形成する。この前縁渦が翼面上に留まっている短い時間は、そのまわりの流れも翼から剥がれず、大きな揚力が発生する。しかし、やがて前縁渦は後方に流れ去ってしまい、流れが翼から剥がれて揚力は小さくなる(図2左上)。ところが小さな翼が回転する(羽ばたく)ときは、前縁渦がらせん状になって翼面上に長く留まり、そのまわりの流れは翼から剥がれず、大きな揚力が維持される(図2右下)[16](Sane, 2003)。
ノドアカハチドリの羽ばたき翼の流体数値シミュレーション(体重3.4 g、翼長45 mm、羽ばたき回数毎秒42回)では、打ち下ろし中に前縁渦が翼面上に留まって大きな揚力が発生することがわかった。打ち上げ中も、打ち下ろし中ほど顕著ではないが、前縁渦が発生して揚力が生まれた。結果として、打ち下ろし中の空気力の鉛直上向き成分は、打ち上げ中の約2.5倍だった。打ち下ろしの方が打ち上げよりも大きな鉛直上向きの力を発生した理由としては、打ち下ろしの方が羽ばたき速度と迎え角が大きく、翼面積もわずかに大きいこと、打ち下ろしでは抗力も鉛直上向き成分を持つこと、などがあげられる[15](Song et al., 2014)。
多様で俊敏な飛行
ハチドリはホバリングが得意なだけでなく、翼の動かし方をさまざまに変えることで、多様で俊敏な飛行が可能である。その優れた飛行能力の一端が垣間見える研究をいくつか紹介したい。
ハチドリに重りをつけて、どれほどの重さを持ちあげてホバリングできるのか実験で調べられた[8](Chai and Millard, 1997)。大型のアオノドハチドリ(体重約7 g)とルリノドシロメジリハチドリ(体重約8 g)は、最大で体重の約2倍の重りを短時間(約0.4秒間)ながら持ちあげて浮上した。
体重約3グラムの小型のアカフトオハチドリとノドグロハチドリの場合は、最大で体重と同程度の重りを約0.6秒間持ちあげた。最大の重りを持ちあげるときは、重りがないときにくらべて、一秒間の羽ばたき回数が約17% から33% 増加し、羽ばたきの振幅も約13% から29% 増加した。これは、ハチドリは羽ばたきの速さと振幅を増やすことで、ホバリング状態から瞬間的に体重の3倍から4倍の空気力を発生して加速急上昇できることを意味する。
前進飛行の最高速度は、風洞(人工的に空気の流れを発生する装置)のなかで、体重約4gのノドアカハチドリを飛行させて計測された[17](Chai and Dudley, 1999)。その結果、最高速度の平均値は秒速14.4m(時速52km)だった。換羽中(風切羽の生え変わりの時期)は、秒速12.4m(時速45km)と少し遅くなることもわかった。
アカフトオハチドリでの同様の実験[10](Tobalske et al., 2007)では、秒速12m(時速43km)まで飛行できた。このときの体の姿勢は、ホバリング中は体軸が水平から50°の角度で前傾して立ち、速度が速くなるほど前傾が深くなり、最高速ではほぼ水平(水平からの角度13°)になった。逆に、横から見た翼端の軌跡は、ホバリング中は水平から15°だったのが、速度が速くなるほど立ち上がり、最高速では水平から68°に立ち上がった(図3)。なお、「ハチドリの翼端は8の字(∞)を描く」としばしば言われるが、図3に見られるように、必ずしも8の字は描かない。
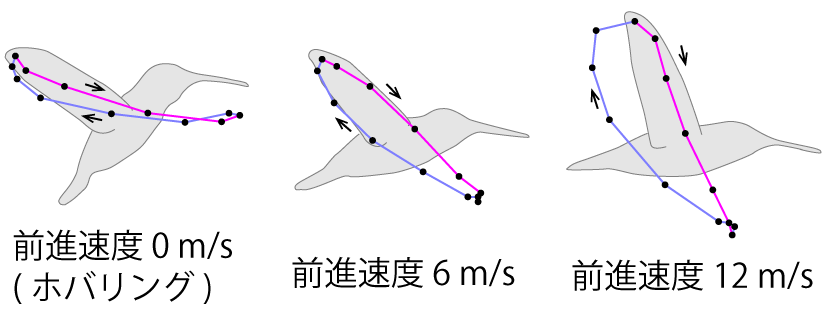
図3 アカフトオハチドリの横から見た翼端の軌跡(文献[10]を基に作成)
ハチドリは真後ろにも飛行できる。風洞内でホバリングしながら蜜を吸う体重約4.6gのアンナハチドリAnna’s Hummingbird の背中から風を当てて、後退飛行を再現した[18](Sapir and Dudley, 2012)。その結果、最大で秒速4.5m(時速16km)で後退飛行できた。このとき、前進飛行とは逆に、体軸は立ち上がり(水平から71°)、羽ばたく方向も後傾して水平になった。つまり、前進や後退は、体軸と翼端軌跡の前傾や後傾で実現される。
またハチドリは気流の乱れにも対応できる。風洞内でホバリングするアンナハチドリに前方から風を当て、上流側には円柱を置いて風を乱したところ、左右の翼の動きを非対称に臨機応変に変えながら、尾羽も使って風洞内での位置を保持した[19](Ortega-Jimenez et al., 2014)。
さらに、ホバリングするアンナハチドリは、壁に開いた幅の狭い穴を、羽ばたき振幅を小さくしつつ横にスライド移動して通過できる。穴をさらに小さくすると、前進飛行して体を深く前傾させ、両翼を後方に引いて羽ばたかず弾丸のように穴を通過する[20](Badger et al., 2023)。
ホバリングして蜜を吸うハチドリを、前方から黒い板を近づけて驚かせると、急反転して後方に飛び去る。この急反転は、わずか5羽ばたき程度での「ピッチアップ&ロール」運動で実現される[21](Cheng et al., 2016)。ピッチアップとは、頭を上げるような体の回転であり、ロールは、体軸まわりの回転である。最初の2羽ばたき程度(約0.06秒間)でピッチアップして体が後方にひっくり返り、続く3羽ばたき程度(約0.09秒間)でロールして上を向いていた腹が下向きに戻り、方向転換が完了する。その間、わずか0.15秒の早業である。
なお、黒い板が動き出してからハチドリが反応するまでの時間は、実験した4種(アオノドハチドリ、ルリノドシロメジリハチドリ、ノドグロハチドリ、アカアシハチドリBroad-billed Hummingbird)のうち3種(アカアシハチドリ以外)でわずか0.03秒未満であり、これはハエやガの視覚からの反応速度である約0.05秒よりも早い。残りの1種も約0.05秒の遅れで、昆虫の反応速度に匹敵する。この早い反応速度もハチドリの俊敏性を支えている。
以上のように、ハチドリは小さな翼を高速で羽ばたかせながらさまざまに動かして、ホバリングと俊敏な飛行を実現している。その飛行能力の限界は未だ明らかではなくさらなる研究が進行中である。本稿では、信頼できるデータに基づく説明を心がけ、多くの研究論文を引用した。これらの論文の中にはインターネット上で無料閲覧できるものがあり、動画が付いているものもある。参考文献リストの該当論文には(無料・動画)と付記した。興味を持たれた方は、ご自身で論文を開きハチドリ研究の世界を覗き見ていただければ幸いである。(たなか・ひろと 情報理工学博士)
参考文献
[1] Gill, F., Donsker, D. and Rasmussen, P., eds. (2024). “Hummingbirds” IOC World Bird List. V14.2.
https://www.worldbirdnames.org/ (ウェブサイト)
[2] Stiles, F. G. (1995). Behavioral, Ecological and Morphological Correlates of Foraging for Arthropods by the Hummingbirds of a Tropical Wet Forest. The Condor 97, 853–878.
https://doi.org/10.2307/1369527
[3] Hummingbird Central.
https://www.hummingbirdcentral.com/hummingbird-migration.htm (ウェブサイト)
[4] Rappole, J. H. and Schuchmann, K.-L. (2003). Ecology and Evolution of Hummingbird Population Movements and Migration.
https://doi.org/10.1007/978-3-662-05957-9_3
[5] Hargrove, J. L. (2005). Adipose energy stores, physical work, and the metabolic syndrome: lessons from hummingbirds. Nutrition Journal 4.
https://doi.org/10.1186/1475-2891-4-36 (無料)
[6] Lawrie, Y., Shannon, T. J., Kirkconnell, A., Clark, C. J., Collinson, J. M. and Kirwan, G. M. (2021). Systematic placement of the bee hummingbird (Mellisuga helenae) (Aves: Trochilidae) and potential consequences for nomenclature of the Mellisugini. Ornitologia Neotropical 32, 45–50.
https://doi.org/10.58843/ornneo.v32i1.947
[7] Fernandez, M. J., Dudley, R. and Bozinovic, F. (2011). Comparative Energetics of the Giant Hummingbird (Patagona gigas). Physiological and Biochemical Zoology 84, 333–340.
https://doi.org/10.1086/660084
[8] Chai, P. and Millard, D. (1997). Flight and size constraints: Hovering performance of large hummingbirds under maximal loading. Journal of Experimental Biology 200, 2757–2763.
https://doi.org/10.1242/jeb.200.21.2757 (無料)
[9] Fernandez, M. J. (2010). Flight Performance and Comparative Energetics of the Giant Andean Hummingbird, PhD thesis, UC Berkeley.
https://escholarship.org/ (無料)
[10] Tobalske, B. W., Warrick, D. R., Clark, C. J., Powers, D. R., Hedrick, T. L., Hyder, G. A. and Biewener, A. A. (2007). Three-dimensional kinematics of hummingbird flight. Journal of Experimental Biology 210, 2368–2382.
https://doi.org/10.1242/jeb.005686 (無料)
[11] Warrick, D., Hedrick, T., Fernández, M. J., Tobalske, B. and Biewener, A. (2012). Hummingbird flight. Current Biology 22, R472–R477.
https://doi.org/10.1016/j.cub.2012.04.057 (無料)
[12] Deeming, D. C. (2023). Allometry of the pectoral flight muscles in birds. Journal of Zoology 319, 264–273.
https://doi.org/10.1111/jzo.13043 (無料)
[13] Kruyt, J. W., Quicazán-Rubio, E. M., van Heijst, G. F., Altshuler, D. L. and Lentink, D. (2014). Hummingbird wing efficacy depends on aspect ratio. Journal of the Royal Society Interface 11.
https://doi.org/10.1098/rsif.2014.0585 (無料)
[14] Maeda, M., Nakata, T., Kitamura, I., Tanaka, H. and Liu, H. (2017). Quantifying the dynamic wing morphing of hovering hummingbird. Royal Society Open Science 4.
https://doi.org/10.1098/rsos.170307 (無料・動画)
[15] Song, J., Luo, H. and Hedrick, T. L. (2014). Three-dimensional flow and lift characteristics of a hovering hummingbird. Journal of the Royal Society Interface 11.
https://doi.org/10.1098/rsif.2014.0541 (無料・動画)
[16] Sane, S. P. (2003). The aerodynamics of insect flight. Journal of Experimental Biology 206, 4191–4208.
https://doi.org/10.1242/jeb.00663 (無料)
[17] Chai, P. and Dudley, R. (1999). Maximum flight performance of hummingbirds. American Naturalist 153, 398–411.
https://doi.org/10.1086/303179
[18] Sapir, N. and Dudley, R. (2012). Backward flight in hummingbirds. Journal of Experimental Biology 215, 3603–3611.
https://doi.org/10.1242/jeb.073114 (無料・動画)
[19] Ortega-Jimenez, V. M., Sapir, N., Wolf, M., Variano, E. A. and Dudley, R. (2014). Into turbulent air. Proc. Royal Society B 281.
https://doi.org/10.1098/rspb.2014.0180 (無料・動画)
[20] Badger, M. A., McClain, K., Smiley, A., Ye, J. and Dudley, R. (2023). Sideways maneuvers in hummingbirds. Journal of Experimental Biology 226.
https://doi.org/10.1242/jeb.245643 (無料・動画)
[21] Cheng, B., Tobalske, B. W., Powers, D. R., Hedrick, T. L., Wethington, S. M., Chiu, G. T. C. and Deng, X. Y. (2016). Flight mechanics of escape manoeuvres. Journal of Experimental Biology 219, 3518–3531.
https://doi.org/10.1242/jeb.137539 (無料)